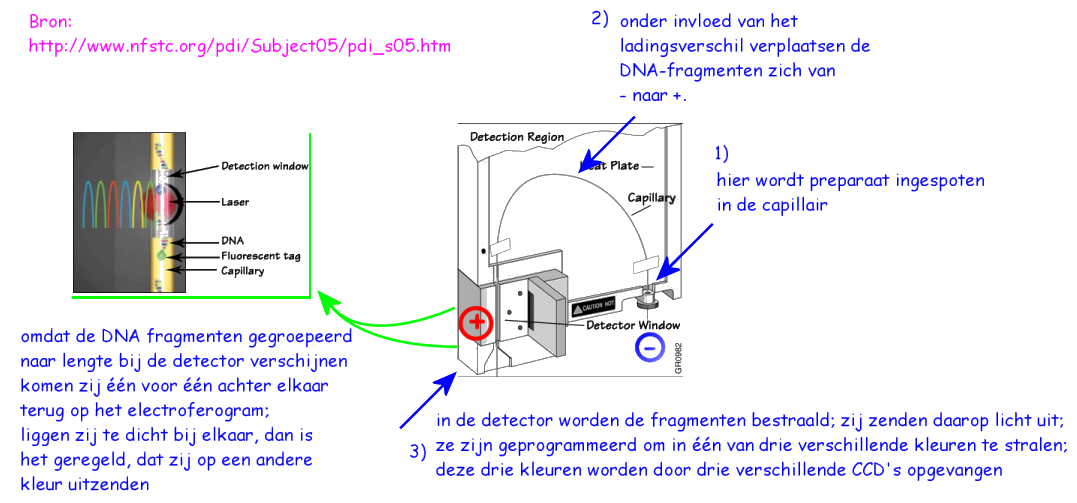
Electroferogram
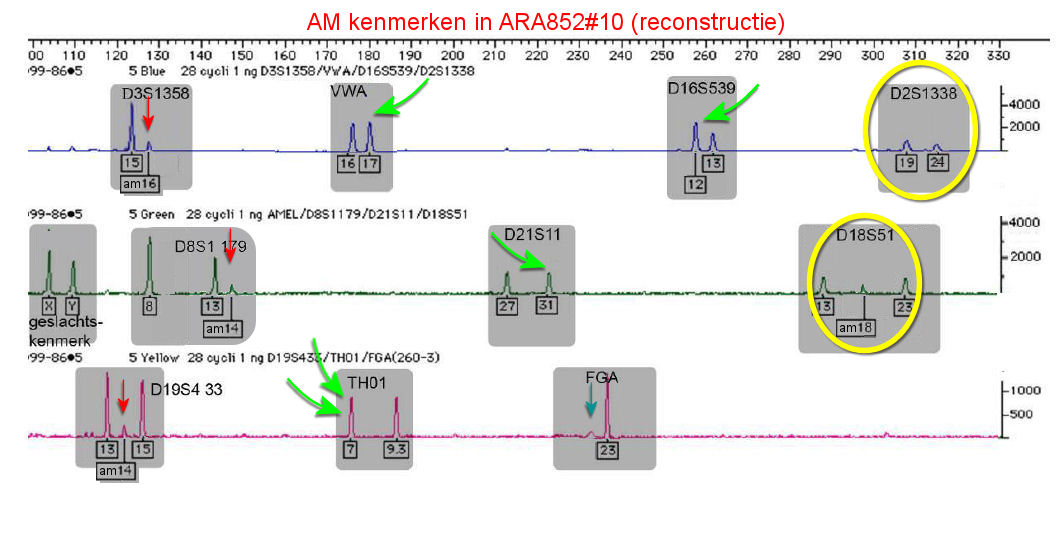
Het electroferogram, dat in ARA852#10 is aangetroffen
moet er zo uitzien:
|
|
|
Reconstructie (*) van het
DNA-profiel, verkregen uit vlek#10. In dit profiel komen
vier kenmerken voor, die EL deelt met vijf kenmerken van
het slo (groene pijlen). Er zijn echter ook een aantal
piekjes, die niet in het profiel van EL thuis horen. Vier
dragen het kenmerk "am". Drie hiervan passen
precies in het
profiel van het slo. Deze werden door de NFI-deskundige
D.A. Kloosterman van het kenmerk "am" voorzien (added
manually), doch hij verzuimde hierbij ook de code "W" te
vermelden, om aan te geven, dat zij matchen met het
profiel van de weduwe. Het vierde hier aangegeven extra
piekje kreeg geen nadere aanduiding, maar viel wel samen
met het profiel van W (zie verderop).
De gele cirkels duiden sporen
aan, waarin geen bijdrage van het slachtoffer is gevonden.
De mogelijke reden hiervan wordt in deze paragraaf
uiteengezet.
* naar Gilder in Derksen
2011 (Leugens over Louwes), pagina 306
|
Hier valt dus op dat er drie/vier piekjes in het profiel
van het slachtoffer vallen en niet in het profiel van EL. Omtrent die drie
piekjes zal zijn geredeneerd, dat het er maar drie zijn, waar zijn dan de
overige piekjes uit het profiel van het slachtoffer? Ofwel, die drie
piekjes zijn 'weggestreept' als toevalligheden.
Maar klopt dit wel? Waar zouden die andere piekjes hebben moeten liggen?
Nu komt de aap uit de mouw:
-
De gevonden am-piekjes liggen allemaal links, dus
betreft allemaal piekjes, die als eerste bij de analyse verschenen in
de zogenaamde detector.
Daarnaast geldt voor de kenmerken D3-16(o) en D8-14
van de weduwe, dat ze "homozygoot" zijn, dus twee gelijk in beide
chromosomen en daardoor extra goed zichtbaar. D19-14 en D19-14.2
liggen onderling op slechts "halve" afstand, en kunnen elkaar
versterkt hebben, gezien de lage resolutie.
-
Een groot aantal van de te verwachten volgende
piekjes van het slachtoffer bevinden zich onder
de reeds bestaande pieken van EL en kunnen daar onmogelijk worden
gedetecteerd. Immers, op de met groen aangewezen plaatsen, hebben EL
en het slachtoffer gemeenschappelijke kenmerken liggen.
-
Voorts zouden zij soms ook heel dicht bij pieken van
EL liggen, waardoor zij de kans lopen te worden aangezien voor
zogenaamde 'artefacten', dwz. signalen die ontstaan door storingen in
de detectie, willekeurige piekjes door schommelingen in de omgeving
(elektrische storingen bijvoorbeeld), 'pull-ups', waarbij een piek
'doorschijnt ' in het verkeerde kleurkanaal en 'stutters', die kunnen
ontstaan doordat het PCR-proces soms een rijtje basen verliest tijdens
het kopiëren.
Dit geldt voor het kenmerk D16-11, dat precies de
stutter-positie voor D16-12 inneemt. Voor D2-23 geldt hetzelfde ten
opzichte van de piek van Louwes D2-24.
-
Dit zal vermoedelijk gelden voor de loci
(aangeduid met de grijze velden), waarin ook de gemeenschappelijke
kenmerken liggen (dus aangeduid met groen). Zie de appendix
onderaan.
-
Daarnaast zien wij in het electroferogram een trend,
die wij veel vaker zullen zien bij het systematisch bekijken van
andere electroferogrammen: naar rechts toe wordt de piekhoogte
gemiddeld fors veel kleiner. Dus is de kans om 'kopje onder te gaan'
in het geweld van artefacten, achtergrond en wat dies meer zijn nog
groter. Let op, hoe laag de pieken zijn in de gele ovalen,
vergeleken met zijn pieken links.
(o)De aanduiding van piek D3S1358-16
kort men af tot D3-16 enzovoort.
Om dit laatste te verduidelijken, kijken wij eerst naar
de techniek van het maken van zo'n electroferogram (Eng.:
electropherogram).
Nadat via PCR (paragraaf 6.2.2) de diafragmenten die
onderzocht worden bij forensische analyse zijn geselecteerd en
vermeerderd, wordt het ontstane preparaat in de capillair van het
geautomatiseerde electroforese-apparaat gespoten:
De verschillende onderdelen van zo'n monster zijn dus
niet allemaal even lang onderweg. De langzaamste fragmenten zijn ongeveer
2x zo lang onderweg, dan de kortste. Dat betekent, dat zij tweemaal zo
lang de tijd hebben, om zich door temperatuursbewegingen over de naaste
omgeving te verspreiden. Voorts kruipt zo'n latere piek veel langzamer
langs de detector, want zijn snelheid is navenant lager. Dan gebeurt er
iets vervelends: de piek van zo'n fragment wordt onscherper; breder en dus
lager:
|
|
|
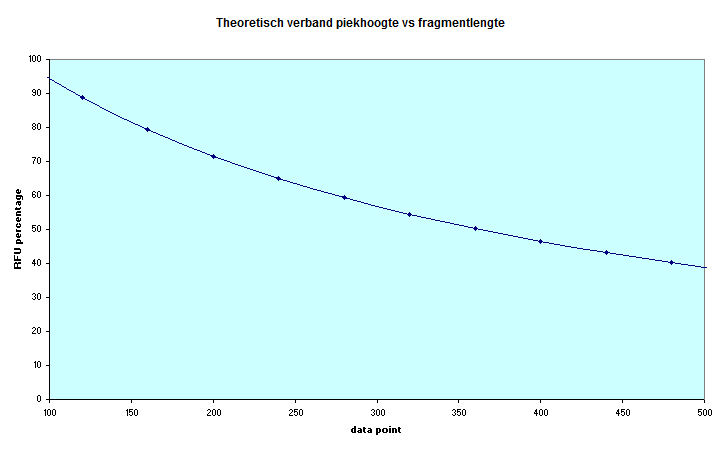
Uitgaande van een omgekeerd
evenredig verband tussen piekhoogte en migration time op
grond van de besproken factoren als pieksnelheid en
diffusie kan de evolutie van de piekhoogte worden
voorspeld.
|
Maar voordien hebben zich nog twee andere
problemen voorgedaan:
- Tijdens de PCR zullen niet alle kopieer-reacties onmiddellijk starten.
Het opstarten van reacties is een stochastisch (= op toevalligheden
berustend) proces, waardoor het mogelijk is, dat bepaalde kopieeracties
niet afgerond worden binnen de daarvoor beschikbare tijd; het PCR proces
is een proces, dat wordt gedicteerd door een op- en neergaande
temperatuur beweging. Het gevolg hiervan is, dat langere fragmenten meer
kans bieden op de mogelijkheid, dat het kopieerproces niet is afgerond
en een fragment wordt geproduceerd, dat deel wordt van de
achtergrondruis.
- tijdens de gebruikte injectie-methode (elektrokinetische injectie)
worden vooral de meest beweeglijke deeltjes opgenomen in de capillair.
Dus eigenlijk een soort electroforese, om de electroforese te kunnen
beginnen. Dit proces trekt de kleine deeltjes voor. Ook hierdoor worden
de pieken aan de linker kant van het uiteindelijk gevormde
electroferogram "bevoordeeld".
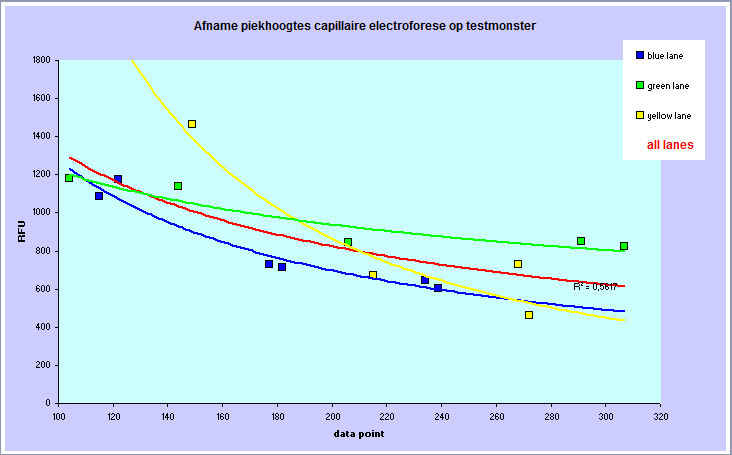
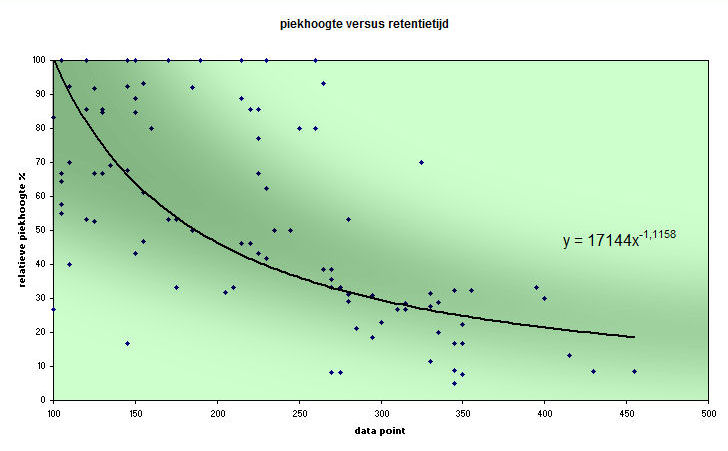
Het gevolg is een afnemende piekhoogte naar rechts in
een electroferogram. Ik heb er een aantal bekeken en de resultaten hiervan
in een diagram weergegeven - het effect is dramatisch:
|
|
|
Piekhoogten in samenhang met
plaatsing van de pieken in een aantal willekeurige
electroferogrammen. Onder aan de pagina is een soortgelijk
diagram van een testmonster opgenomen met eenzelfde
resultaat.
|
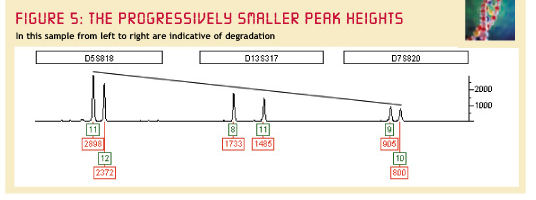
Dit alles blijkt al eerder geconstateerd, in http://www.bioforensics.com/articles/champion1/champion1.html:
|
|
|
Degeneratie ter verklaring.
Hier wordt hetzelfde effect gesignaleerd. De
piekafname wordt hier verder geaccentueerd, doordat de
pieken aan de rechter zijde corresponderen met langere
DNA-fragmenten, voordat deze werden gekopieerd. Langere
DNA-fragmenten zijn op basis van pure kansrekening meer
vatbaar voor degeneratie. In de DMZ is dit een belangrijke
factor (4 jaar!).
Zie appendix voor resultaten, waarbij degeneratie is
uitgesloten.
|
Samen met de eerder genoemde factoren, is hiermee het
fenomeen afdoende verklaard.
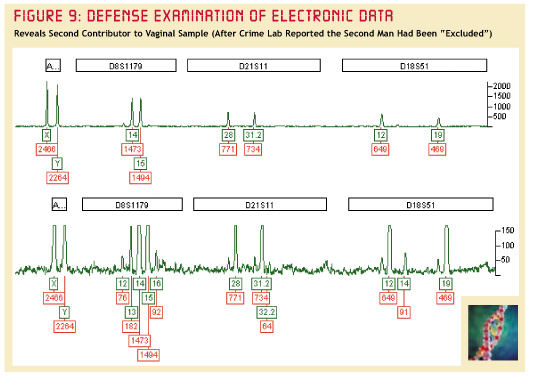
In dit artikel wordt een casus opgevoerd, die als twee
druppels water lijkt op de casus van vlekje #10:
|
|
|
Doordat de ene bijdrage van
het DNA-spoor zulke hoge pieken veroorzaakte, werd de
tweede bijdrage gemaskeerd. In het geval van vlekje#10
doet zich precies hetzelfde voor. De pieken in vlekje #10
zijn erg sterk.
D8S1179-13 en D8S1179-16
zijn met een beetje goede wil nog in het bovenste diagram
zichtbaar. D21811-32.2 is echt niet te zien daar. In
D8S1179 is nu ook het DNA van het slachtoffer zichtbaar.
|
Zie, hoe sterk ook in het getoonde deel-electroferogram
de piekhoogte naar rechts afloopt!
Het fenomeen staat bekend onder twee namen, de
ski-helling en de degradatie-helling. In deze uiteenzetting nogmaals de
oorzaken: A ski slope is not necessarily a degradation slope.pdf
Een concreet voorbeeld
Hoe pakt dit nu uit in de praktijk? In de
volgende illustratie is het electroferogram van spoor #10 herhaald, nu
onmiddellijk gevolgd door het diagram van spoor #6. Spoor #6 is in zijn
totaliteit alleen toegeschreven aan het slachtoffer. Het is incompleet,
een incompleet profiel kan dus ook. In spoor #6 heb ik met verschillende
kleurcodes een aantal pieken aangeduid, waarvan ik in spoor #10 heb
aangeduid, hoe het spoor zich daar zou manifesteren, indien spoor #10 een
mengprofiel zou zijn van Louwes en van spoor #6.
|

|
| Onder: spoor #6. De rood
aangeduide sporen zijn ook in spoor #10 (boven)zichtbaar. De
groen aangeduidde sporen vallen in spoor #10 samen met daar
aanwezige pieken van Louwes. De blauw aangeduidde pieken
vallen opposities, die in spoor #10 ook als stutterpieken
kunnen worden geduid. In zo'n geval is een specifieke
toewijzing aan de één of de ander nietmogelijk. De bruin
aangeduide pieken staan op posities, die in diagram #10
onmiddellijk zouden opvallen. Maar zijn daarin niet
gesignaleerd. Een aantal
van deze pieken zijn ook in spoor #6 niet gesignaleerd.
Terwijlspoor #6 toch echt aan het slachtoffer werd
toegeschreven. Omgekeerd kunnen een aantal - blauwe -
pieken, die hier ontbreken gewoon in spoor#10 staan, zonder
op te vallen. De redenen van ontbreken van
dergelijke pieken staan hierboven beschreven. |
Nog een voorbeeld
Hieronder staan twee electroferogrammen,
nu gewoon als enkele diagrammen op een enkele x-as, dus alle pieken gewoon
achter elkaar. Het bovenste diagram toont alle pieken in spoor #10. Het
onderste diagram toont het referentiespoor van Louwes, gemaakt op basis
van zijn wangslijmvlies met daardoor gemengd 5% van het
referentie(bloed-)spoor van het slachtoffer. Zoek de verschillen.
|
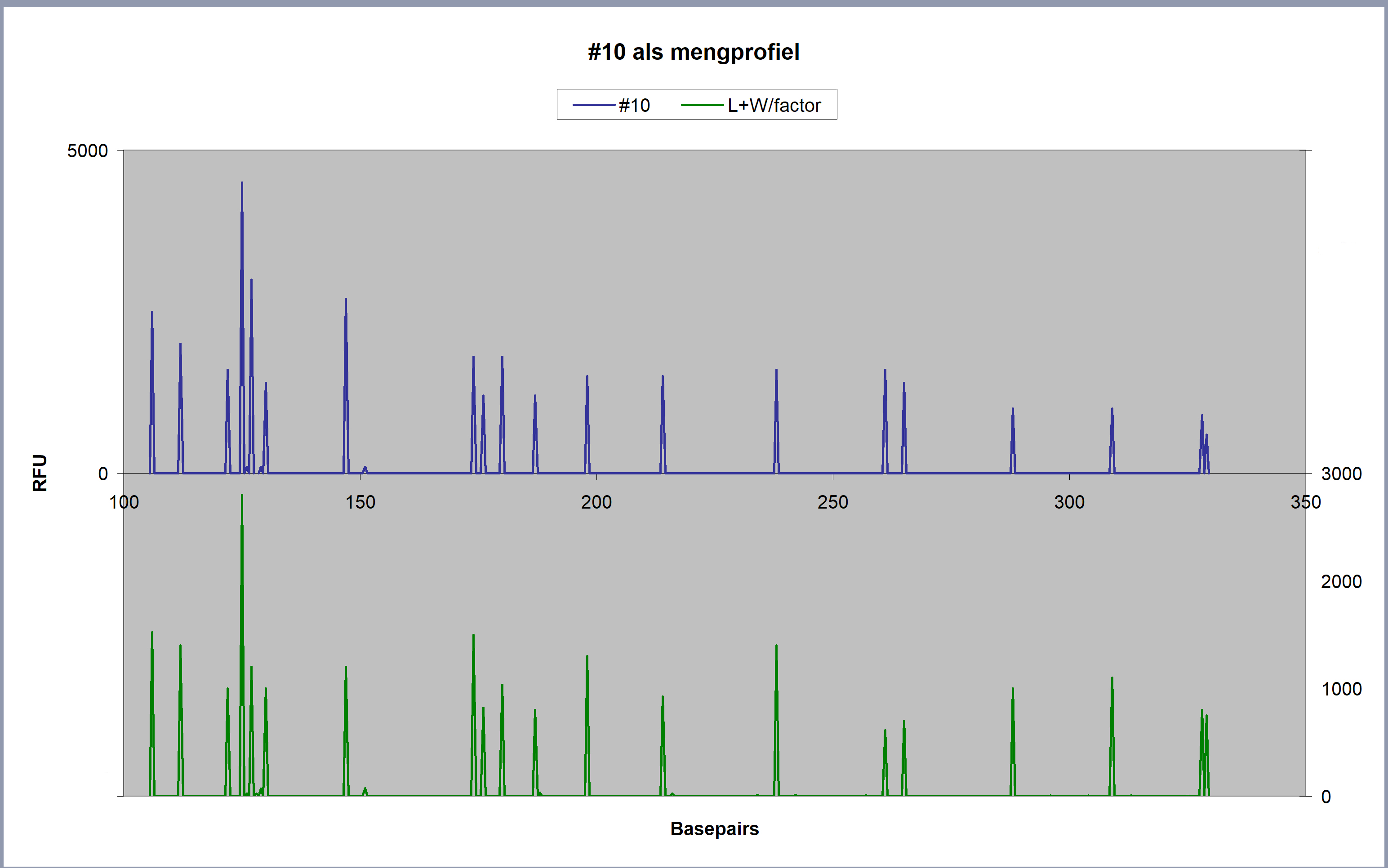
|
Boven: De pieken in spoor
#10. Onder: De pieken in het referentiespoor van Louwes,
gemengd met het refentiespoor van het slachtoffer, waarvan een
5%-bijdrage is toegevoegd (in overeenstemming met de aanname,
dat het bloedspoor in spoor #10 van het slachtoffer afkomstig
is, zie de paragrafen 6.3 [rekenen]
en 6.3.2. [vlek
#10 uitgerekend]) Vanaf 200 basepairs duiken de pieken
van het slachtoffer onder het detectieniveau van 50 RFU, ze
verdrinken daar in de ruis van de basislijn.
|
Conclusie:
Er moet beter gekeken worden. Dan zal blijken -in zover
dat eigenlijk niet al gebleken is, dat er TWEE DNA-bijdragen in spoor
ARA852#10 aanwezig zijn, een sterk (primair) spoor en
een zwak (secundair) spoor:
-
Het sterke spoor is ontegenzeggelijk van
EL. Het kan veroorzaakt zijn door speeksel, zoals zoveel van zijn
sporen wel moeten zijn veroorzaakt door speeksel, gezien het ontbreken
van andere zichtbare aanwijzingen.
-
Het zwakke spoor is nu al via drie
markers verbonden met het slachtoffer. Van het slachtoffer zijn rond
de 20 bloedsporen aangetroffen, die met slechts heel weinig - of
helemaal zonder- markers aan haar gekoppeld kunnen worden.
NIETS weerspreekt de stelling, dat het zichtbare
bloedspoor in het uitgeknipte monster ARA852#10 de leverancier van het secundaire
spoor is. Het zou de te falsificeren hypothese moeten zijn.
Appendix:
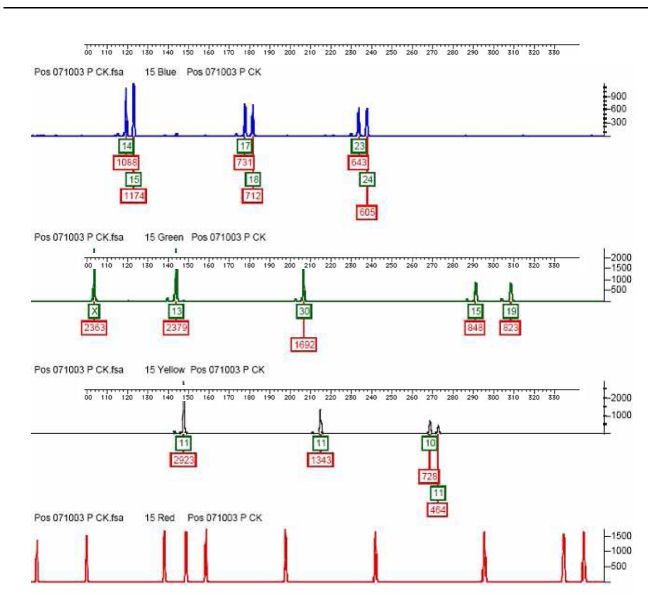
Afname piekhoogtes in een testmonster (9947A positif
control sample; dus degeneratie uitgesloten):
Stutters zijn heel effectief in het 'verdonkeremanen' van een tweede
profiel:
 |
 |
|
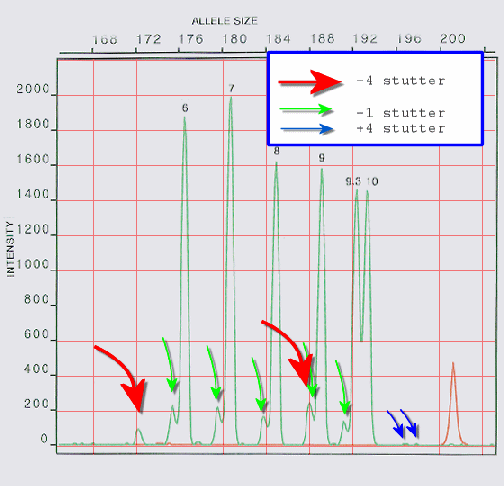
Electroferogram van een
'ladder' (groep DNA-fragmenten t.b.v. ijking) op de ABI
PRISM 310. Opmerkelijk zijn de regelmatige afname van de
piekhoogten en het optreden van duidelijke stutters. De
meeste -4-stutters zijn onzichtbaar onder de naburige
pieken. De -4-stutter van allel 9.3 zit onder de
-1-stutter van allel 9 (die daardoor hoger is).
|
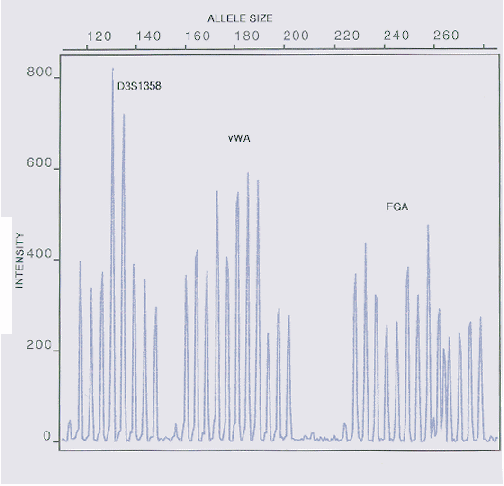
Nog een analyse uit dezelfde
bron, maar nu met een wat bredere schaal, waardoor de
afname in piekhoogte nog verder is geaccentueerd. In
dergelijke gevallen (meting van een ijkmengsel) is
degradatie uitgesloten. Dezelfde stutterpatronen zijn ook
hier zichtbaar.
|
|
Zie: http://www.promega.com/geneticidproc/ussymp7proc/0717.html
(1996).
|